Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Кинетика процессов утилизации субстрата, образования продуктов метаболизма и биомассы в культурах клеток
Кинетика образования продуктов метаболизма
Химически структурированные модели кинетики образования продуктов жизнедеятельности клеток
В сравнении с изучением кинетики роста индивидуальной клетки и популяций клеток в проблеме кинетики процессов образования продуктов метаболизма разработке структурированных моделей посвящено относительно небольшое число работ. Можно ожидать, однако, что развитие структурированных моделей клеточного роста в ближайшем будущем потребует разработки ряда таких моделей. В этом разделе мы рассмотрим интересный процесс образования одного из множества вторичных метаболитов, на примере которого познакомимся с рядом важных аспектов процессов микробиологического синтеза антибиотиков, а также с новыми подходами к моделированию кинетики таких процессов.
Синтез ряда микробных антибиотиков и других продуктов вторичного метаболизма ингибируется высокими концентрациями фосфатного иона. Поскольку для роста клеток фосфат совершенно необходим, очевидно, должен существовать некоторый оптимальный уровень его концентрации. Действительно, такой оптимальный уровень начальной концентрации фосфата наблюдался, например, при биосинтезе алкалоидов Claviseps purpurea (рис. 7.32).
Приведенные на рис. 7.32 экспериментальные данные хорошо моделируются следующими уравнениями кинетики клеточного роста периодической культуры и образования продукта жизнедеятельности [26].
Рост клеток:
![]()
Концентрация фосфата в среде:
![]()
Концентрация фосфата в клетке:
![]()
Образование алкалоидов:
![]()
В этой модели удельная скорость роста описывается уравнением Теиссье, в котором одной из независимых переменных является внутриклеточная концентрация фосфата рi. (Эта переменная равна числу граммов КН2РО4, содержащемуся в 1 г биомассы, и отражает состав клетки. Все другие переменные этой модели, описывающие компоненты системы, измеряются в граммах на 1 л культуры.) Выражение k2x2 отражает скорость лизиса клеток.
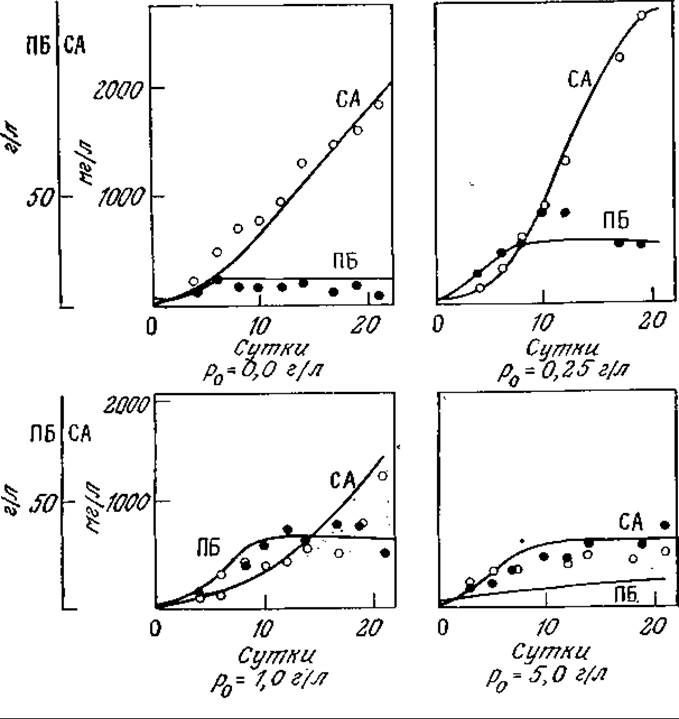
РИС. 7.32. Изменение во времени плотности биомассы (ПБ) С. purpurea (●, г сухого вещества в 1 л) и суммы продуцируемых алкалоидов (СА) (О, мг/л) при различных начальных концентрациях фосфата. Кривые отражают соответствующие расчетные данные, полученные с помощью описанных в тексте математических моделей. [Воспроизведено с разрешения из статьи: Раzоutova S., Votruba J., Rehäcek Z., A Mathematical Model of Growth and Alkaloid Production in the Submerged Culture of Claviceps purpurea; Biotech. Bioeng., 23, 2837 (1981).]
В результате лизиса фосфат выделяется в среду; как показывает второе слагаемое правой части уравнения (7.95), его количество пропорционально содержанию фосфата в клеточной массе Yp/х и внутриклеточной концентрации фосфата рi. Первое слагаемое той же части того же уравнения описывает активный транспорт фосфата в клетку; кинетика этого процесса характеризуется наличием режима насыщения. Уравнение материального баланса по внутриклеточному фосфату [уравнение (7.96)] является вариантом уравнения (7.67); здесь учтено, что скорость его образования равна разности между скоростью транспорта фосфата в клетку и скоростью включения фосфата в компоненты клетки. В уравнении скорости образования продуктов принято во внимание ингибирование процесса фосфатом в том виде, в каком этот эффект учитывался ранее при моделировании репрессии фермента фосфатазы.
Таблица 7.5. Значения параметров в структурированной модели кинетики биосинтеза алкалоидов в С. purpureaа
Параметр |
Размерность |
Значение |
Соответствующий доверительный предел |
k1 |
сутки-1 |
0,5 |
0,058 |
k2 |
л/г в сутки |
0,016 |
0,0024 |
k3 |
сутки-1 |
0,0575 |
0,017 |
k4 |
мг/г в сутки |
6,028 |
1,69 |
Yр/x |
— |
0,0025 |
0,0024 |
K1 |
— |
1,87∙10-4 |
1,73∙10-4 |
К2 |
г/л |
4,29∙10-4 |
6,67∙10-4 |
К3 |
— |
4,65∙10-4 |
9,84∙10-5 |
а Воспроизведено с разрешения из статьи: Pazoutova S., Voriruba Rehäcek Z., A Mathematical Model of Growth and Alkaloid Production in the Submerged Culture of Claviceps purpurea, Biotech. Bioeng., 23, 2837 (1981).
Сплошные кривые на рис. 7.32 были вычислены с помощью описанной модели, а значения соответствующих параметров перечислены в табл. 7.5. Эта структурированная модель хорошо отражает закономерности изменения биомассы и массы алкалоидов во времени, в том числе наличие максимума на кривой накопления суммы алкалоидов, но в то же время при высоких концентрациях фосфата ее точность недостаточно высока. С помощью такой модели, в частности, было показано, что максимальное количество суммы алкалоидов продуцируется при начальной концентрации фосфата в среде 0,17 г/л. Математическое моделирование различных методик введения фосфата (в том числе добавления фосфата по частям в различные фазы процесса) показало, что таким путем нельзя добиться существенного повышения выхода алкалоидов по сравнению с обычным периодическим процессом при оптимальной начальной концентрации фосфата в среде. В других случаях, которые мы рассмотрим в последующих главах, периодические процессы с добавлением субстрата оказываются очень полезными.