Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Стехиометрия и энергетика метаболических превращений
Живая клетка представляет собой сложный химический реактор, в котором осуществляется более 1000 независимых катализируемых ферментами реакций. Тем не менее биологические системы подчиняются тем же основным законам сохранения вещества и энергии и тем же принципам термодинамики, которые хорошо знакомы нам по процессам химической технологии. В настоящей главе мы будем рассматривать клетку именно как химический реактор, концентрируя внимание на стехиометрических соотношениях, процессах выделения или поглощения энергии, характерных для живых организмов.
Совокупность всех осуществляющихся в клетке химических превращений называется метаболизмом. На рис. 5.1 приведена упрощенная схема некоторых наиболее важных процессов метаболизма в бактерии Escherichia coli. Нетрудно видеть, что метаболические реакции большей частью объединяются в последовательности, называемые путями метаболизма, причем связь между ними осуществляется, во-первых, через циклические, закрытые пути, в которых продукт последней стадии является исходным веществом для первой стадии, и, во-вторых, посредством ответвлений от основных путей, соединяющих одну последовательность реакций с другой. Все стрелки на рис. 5.1 обозначают одну или несколько катализируемых ферментами реакций, в результате которых одни компоненты клетки (метаболиты) превращаются в другие.
Отдельные клетки и их популяции представляют собой более упорядоченную по сравнению с окружающей средой систему, а упорядочение любой системы требует затрат энергии. Сейчас мы уже более или менее детально знаем, каким образом необходимая для этого энергия извлекается из окружающей клетку среды. С другой стороны, — это, вероятно, наиболее важно с точки зрения биохимической технологии, — изучение энергетического обмена помогает понять и объяснить основные различия между функционированием клеток в присутствии и в отсутствие кислорода. Как мы уже отмечали, такие условия называются аэробными и анаэробными соответственно. Одни клетки (облигатные, или строгие, анаэробы) вообще не используют свободный кислород, другим клеткам (облигатным аэробам) кислород необходим, а третьи клетки могут расти как в присутствии кислорода, так и без него. Метаболически универсальную третью группу клеток называют факультативными анаэробами; к ним относятся, в частности, хорошо известные дрожжи.
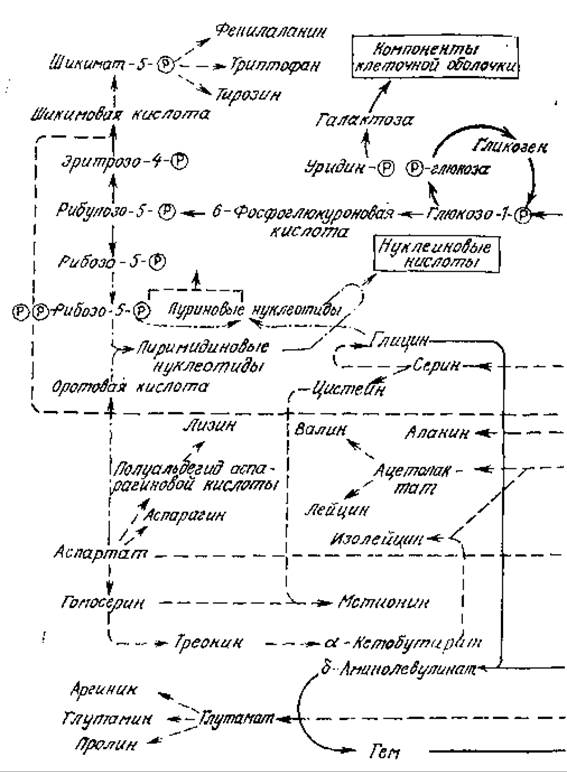
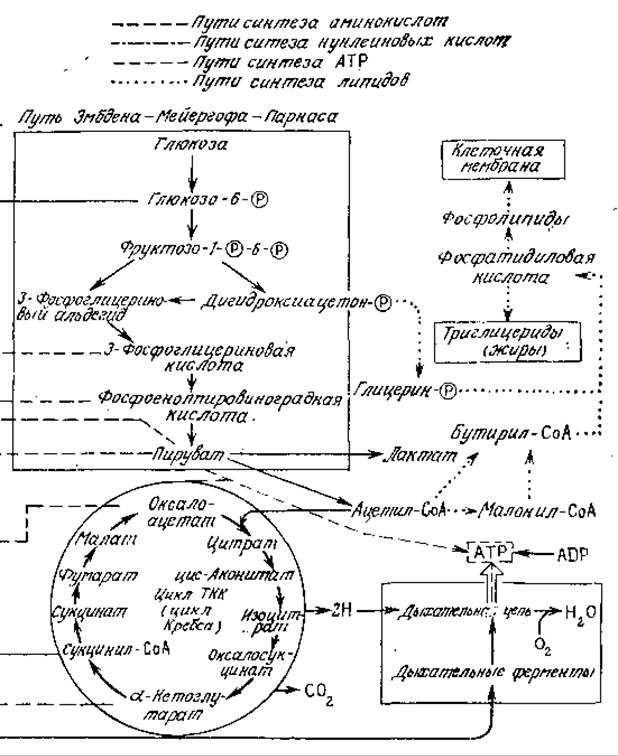
РИС. 5.1. Схематическое изображение некоторых основных путей метаболизма бактерии Е. coli. Дж., Уотсон. Молекулярная биология гена. — М.: Мир, (1978.)
Микроорганизмы усваивают два вида энергии — световую и химическую. Фототрофами называют организмы, для которых основным источником энергии является свет, а хемотрофами — организмы, получающие энергию в результате деградации определенных питательных веществ. В соответствии с природой последних хемотрофы подразделяют далее на ряд подгрупп. Так, литотрофы окисляют неорганические вещества, а органотрофы для извлечения энергии используют органические питательные вещества. Несколько конкретных примеров мы приведем после того, как детальнее ознакомимся с основами питания клеток.
Полученная тем или иным способом энергия обычно хранится и транспортируется (внутри клетки) в виде удобных высокоэнергетических промежуточных соединений типа АТР (разд. 2.3.1). Клетка использует эту энергию для выполнения следующих трех функций: химического синтеза больших или сложных молекул, (т. е. для роста); транспорта ионных и нейтральных веществ в клетку, из клетки, а также в клеточные органоиды и из них; механической работы, связанной с делением и передвижением клеток. Ни один из этих процессов не происходит спонтанно, а их результатом является повышение свободной энергии клетки. По этой причине они осуществляются только совместно с другими реакциями, сопровождающимися большим по абсолютной величине понижением свободной энергии. К этой теме мы вернемся в разд. 5.1.
Эффективность использования свободной энергии в биосинтезе довольно высока и обычно превышает 20%. Транспорт веществ также происходит с участием АТР; этот процесс уникален в том отношении, что ом обеспечивает перенос небольших молекул и ионов через мембраны против градиента концентрации даже в тех случаях, когда отношение концентраций транспортируемого вещества по обе стороны мембраны составляет 105. Механическая работа, очевидно, совершается при делении клеток, а также при движении бактерий и простейших. В мышечной активности животных и движении сперматозоидов также участвует АТР; прямое превращение химической свободной энергии в механическую работу без участия таких посредников, как электричество или горячие газы, представляет собой еще одно уникальное свойство живых систем. Частично теряющаяся в ходе таких процессов химическая энергия превращается в тепловую, что необходимо учитывать в технологических процессах с участием растущих популяций клеток.
Таблица 5.1. Важнейшие источники химических элементов
Химический элемент |
Источник |
Углерод |
СO2, сахара, белки, жиры |
Азот |
Белки, NH3, NO3- |
Сера |
Белки, SO42- |
Фосфор |
PO43- |
Чтобы расти и самовоспроизводиться, клетка должна потреблять вещества, необходимые для построения мембран, белков, хромосом, стенок и других структурных элементов. Из основных химических элементов для жизни необходимы прежде всего углерод, азот, сера и фосфор; водород и кислород могут быть получены из компонентов среды или в некоторых случаях из воды. Типичные источники этих элементов приведены в табл. 5.1, а на рис. 5.2 изображена схема их утилизации некоторой воображаемой бактерией. Этот организм можно было бы назвать гетеротрофом, поскольку он получает необходимый углерод из органического вещества. Другие микроорганизмы могут использовать более простые питательные вещества; так, аутотрофы всю свою потребность в углероде удовлетворяют за счет СО2. Объединив эту градацию с указанной выше классификацией организмов в соответствии с источниками энергии, мы можем определить совокупность признаков, описывающих метаболизм клетки. Соответствующая номенклатура и примеры организмов каждого типа приведены в табл. 5.2. Использование разными клетками различных источников энергии и углерода говорит о том, что и химизм протекающих в них процессов неодинаков. В настоящей главе основное внимание уделено изучению наиболее общих путей метаболизма, характерных для самых разных типов живых клеток; важные проблемы дифференциации путей метаболизма обсудим позднее.
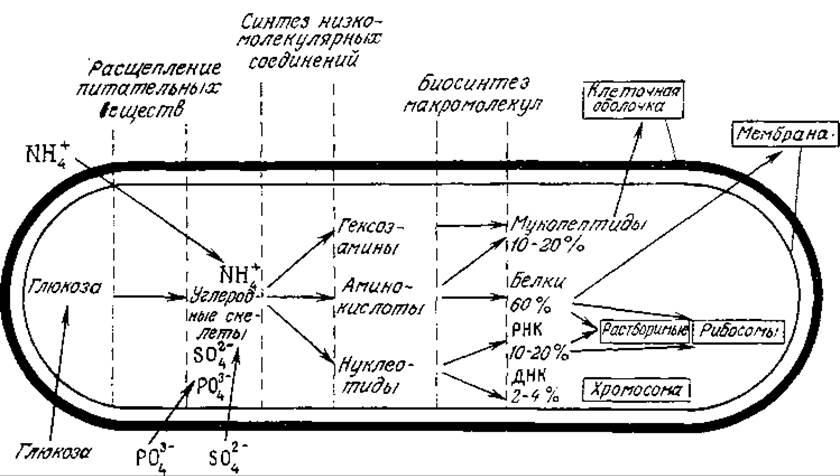
РИС 5.2. Схематическое изображение биосинтеза макромолекулярных соединений бактерией из несложных питательных веществ. [Воспроизведено с разрешения из работы: Biochemistry of Bacterial Growth, Mandelstam J., McQuillen K., (eds.), 2d ed., p. 4, Blackwell Scientific Publications, Oxford.
Таблица 5.2. Классификация и номенклатура организмов в зависимости от источников углерода и энергии
|
Источник энергии |
||
Источник углерода |
Химический |
Световой |
Органические соединения |
Хемогетеротрофы (высшие животные, простейшие, грибы и большинство бактерий) |
Фотогетеротрофы (некоторые бактерии, ряд водорослей- эукариот) |
СO2 |
Хемоаутотрофы (некоторые бактерии) |
Фотоаутотрофы (высшие растения, водоросли-эукариоты, сине-зеленые водоросли и некоторые бактерии) |
На рис. 5.2 происходящие в клетке реакции разделены на три группы: деградация питательных веществ, биосинтез веществ небольшой молекулярной массы и биосинтез полимерных макромолекул. Как мы уже отмечали, каждая реакция катализируется специфическим ферментом; последние, таким образом, выполняют важнейшую функцию, определяя, какие реакции и с какой скоростью будут протекать в клетке.
Чтобы оценить важность регуляции относительных скоростей реакций, полезно взглянуть на химию клетки с другой стороны. Многие находящиеся в клетке вещества (метаболиты) могут одновременно подвергаться действию нескольких ферментов, которые, например, могут окислять, восстанавливать или конденсировать их с другими веществами. В результате происходящие в клетке последовательности реакций пересекаются и перекрываются самым замысловатым образом, как это показано, в частности, на рис. 5.1. Обратите внимание, например, на то, что конечный продукт пути Эмбдена — Мейергофа (называемого также путем Эмбдена — Мейергофа — Парнаса или ЭМП) пируват может далее использоваться в пяти различных путях биосинтеза. Хотя приведенная на рис. 5.1 схема может на первый взгляд показаться очень сложной, нельзя забывать, что па самом деле здесь изображена лишь ничтожная доля из приблизительно 1000 происходящих в клетке реакции. К тому же на рис. 5.1 не указано, что многие стадии этих путей метаболизма обратимы; обратимость реакций обеспечивает возможность как синтеза, так и деградации биоорганических соединений. Несколько позже в этой же главе мы более подробно рассмотрим некоторые из этих путей метаболизма, а также контролирующие и регулирующие их ферментативные механизмы.
На рис. 5.1 и 5.2 отсутствует и другая важная деталь. Мы не показали здесь пути выведения конечных продуктов метаболизма из клетки. Многие из этих веществ не нужны для жизнедеятельности клетки, а другие, например антибиотики или внеклеточные ферменты, выполняют определенную функцию. Некоторые из этих продуктов метаболизма (спирты, органические кислоты, аминокислоты, антибиотики и многие другие) представляют большую ценность для человека, что экономически оправдывает их промышленное производство с помощью выращивания культур клеток. Для таких целей обычно стремятся найти неэффективный в биологическом смысле штамм клеток, который продуцирует интересующее нас вещество в количествах, намного превышающих потребности организма. По этой причине в настоящей главе мы попытаемся также проследить пути взаимопревращений различных химических соединений внутри клетки, с тем чтобы понять, что можно производить с помощью живых организмов. Хотя здесь и в последующих главах мы лишь коснемся этой темы, далее этого поверхностного взгляда будет достаточно, чтобы оценить невероятную сложность и почти неисчерпаемое разнообразие происходящих в биологическом мире химических превращений.
Химический состав микроорганизмов или животных клеток одного типа изменяется в сравнительно узких пределах, и метаболическая активность клетки подчиняется обычным принципам стехиометрии и термодинамики. Так, для синтеза заданного количества клеточного вещества требуется определенное количество углерода, азота и кислорода из непосредственного окружения клетки. Далее, если нам известна химическая форма, в которой эти элементы потребляются клетками из среды, мы можем установить дополнительные ограничения, связывающие количество утилизованных клетками питательных веществ с массой образовавшихся клеток вообще и определенных продуктов биосинтеза в частности. Кроме того, поскольку синтез всех необходимых для построения новых клеток веществ требует затрат энергии, то необходимо учитывать и источники этой энергии. Это в свою очередь налагает соответствующие ограничения на количество используемой клеткой химической или световой энергии, так как клетка усваивает энергию в виде АТР в результате определенной последовательности реакций.
По сути дела, уже одни ограничения материального баланса и принципы термодинамики могут служить достаточной основой для выработки математических выражений, в весьма упрощенном виде описывающих клеточную активность; в этом подходе, рассматриваемом подробнее в разд. 5.10, совершенно не учитываются данные о внутреннем химизме происходящих в клетке процессов, а изучаются только параметры, характеризующие рост клеточной массы, потребление клетками субстратов и продуцирование определенных продуктов метаболизма. Такой подход дает возможность получить уравнения, позволяющие по ряду легко измеряемых величин сделать определенные выводы о некоторых необходимых условиях работы биологического реактора и проверить соответствие экспериментальным данным.
Изучение материала мы начнем с фундаментальных механизмов, лежащих в основе жизнедеятельности клетки, и затем перейдем к конечным результатам процессов, осуществляющихся в соответствии с этими механизмами. Прежде всего более или менее подробно проанализируем роль АТР, универсального переносчика энергии в клетке, а также типы реакций, в которых АТР поглощает и отдает энергию. В этих же разделах мы рассмотрим важные окислительно-восстановительные процессы и способствующие их осуществлению в клетке переносчики электронов (NADH и NADPH), затем перейдем к изучению некоторых путей катаболизма, связанных с разложением питательных веществ (из которых основное внимание будет уделено глюкозе), определим энергетику и стехиометрию этих процессов и вкратце рассмотрим также механизмы фотосинтеза. Далее с той же точки зрения мы изучим некоторые пути биосинтеза различных веществ в клетке, после чего перейдем к рассмотрению взаимосвязи между путями метаболизма, в которых генерируются энергия и электроны, и использующими их последовательностями реакций. Наконец, мы проанализируем общие стехиометрические соотношения, характерные для клеточного роста, и в завершение главы рассмотрим стехиометрию образования продуктов метаболизма и ее связь со стехиометрией клеточного роста.