Основы биохимической инженерии Часть 1 - Бейли Дж., Оллис Д. 1989
Стехиометрия и энергетика метаболических превращений
Катаболизм углерода
Другие пути катаболизма углеводов
Прежде всего рассмотрим две другие последовательности реакций, в которых осуществляется катаболизм глюкозы. Пентозофосфатный цикл или пентозофосфатный путь (называемый также гексозомонофосфатным путем или шунтом) начинается с окисления глюкозофосфата:
Глюкозо-6-фосфат+NАDР+→ 6-фосфоглюконат + NADPH + H+
Основной задачей пентозофосфатного пути является обеспечение клетки NADPH, выполняющим роль переносчика электронов в биосинтетических реакциях. Схема всех происходящих на этом пути реакций довольно сложна (см. стр. 456 в работе [1]), но в целом стехиометрию пентозофосфатного пути можно выразить следующим довольно простым уравнением:
Глюкоза + 12NADP+ + 7H2O + ATP →
→ 6СО2 + Рi + 12 (NADPH + H+) + ADP (5.15)
Таким образом, конечным результатом пентозофосфатного пути является полное окисление глюкозо-6-фосфата до СО2 с передачей всех электронов (Н) NADP.
В указанном пути потребляется АТР. Чтобы обеспечить клетку АТР, пентозофосфатный цикл может быть прерван на стадии, когда общая стехиометрия процесса выражается уравнением
3 Глюкоза + 6NADP+ + АТР →
→ 2 фруктозо-6-фосфат+глицеральдегид-3-фосфат + 3СO2+
+ 6(NADPH + H+) + ADP + Pi (5.16)
Если дальнейшие превращения фруктозо-6-фосфата и глицеральдегид-3-фосфата до пирувата будут осуществляться по пути ЭМП, то на этом пути произойдет и фосфорилирование 6 молекул ADP (в расчете на 3 молекулы глюкозы). В результате из моля глюкозы образуется 5/3 моля АТР, но общий выигрыш в энергии будет все же меньше, чем в метаболическом пути ЭМП.
В то же время пентозофосфатный путь имеет и свои преимущества, обусловленные прежде всего тем обстоятельством, что в нем в качестве промежуточных веществ образуются рибозо-5-фосфат и эритрозо-4-фосфат, являющиеся важными предшественниками в синтезе пуринов и пиримидинов в клетке. В метаболическом пути ЭМП такие соединения не образуются. По этой причине микроорганизмы типа молочнокислых бактерий, располагающие для катаболизма углеводов только путем ЭМП, в анаэробных условиях могут расти только на сложных питательных средах, в состав которых входят пентозы, пурины и пиримидины. Напротив, бактерия Е. coli даже в анаэробных условиях способна расти и на более простых средах, поскольку в ней реализуются одновременно и путь ЭМП, и пентозофосфатный путь (в некоторых условиях, например, 75% углеводов катаболизируется по пути ЭМП, а 25% — по пентозофосфатному пути). Этот факт является иллюстрацией общего принципа, согласно которому клетки могут катаболизировать данное питательное вещество несколькими путями одновременно, что, по всей вероятности, обеспечивает их оптимальный рост за счет удовлетворения потребностей как в энергии, так и в веществах- предшественниках.
Последний из рассматриваемых здесь путей катаболизма глюкозы называется путем Энтнера — Дудорова (ЭД). Общая стехиометрия этой последовательности реакций выглядит следующим образом:
Глюкоза + ATP + NADP+→
→ глицеральдегид-3-фосфат+пировиноградная кислота +
+ ADP + NADPH + H+ (5.17)
Если учесть, что последующее превращение глицеральдегид-3-фосфатд в пируват по той же схеме, что и в пути ЭМП, сопровождается фосфорилированием двух молекул ADP, то становится очевидным, что с энергетической точки зрения путь ЭД, в котором образуется моль АТР из моля глюкозы, не очень выгоден. Прежде чем перейти к изучению катаболизма других питательных веществ, следует подчеркнуть, что описанные выше основные пути катаболизма глюкозы далеко не исчерпывают возможности бактерий.
Ранее мы упоминали, что в качестве источников углерода микроорганизмы могут утилизировать различные соединения. С детальным анализом этой проблемы читатель может ознакомиться в работе [3]; мы же ограничимся несколькими примерами, иллюстрирующими только основные пути катаболизма отличных от глюкозы веществ.
Утилизация других сахаров часто начинается с их превращения с помощью одной или нескольких реакций в глюкозу или в одно из промежуточных соединений в путях ее катаболизма. В качестве одного из примеров на рис. 5.5 приведена последовательность реакций, посредством которых Е. coli начинает ассимиляцию дисахарида лактозы. Особенность этого пути катаболизма заключается в специфическом механизме, обеспечивающем утилизацию галактозы. Обратите внимание на циклический характер утилизации уридиндифосфата (UDP), который выполняет таким образом функции катализатора. В качестве второго примера можно привести утилизацию фруктозы некоторыми бактериями; в этом случае в результате двух катализируемых ферментами реакций фруктоза превращается в соответствующий 1,6-дифосфат, который затем может быть включен в путь ЭМП. Утилизация бактериями пентоз обычно начинается с последовательности реакций, в результате которых образуется ксилулозо-5-фосфат; последний может быть включен в пентозофосфатный путь, а может претерпевать расщепление по фосфокетолазному пути (рис. 5.6).
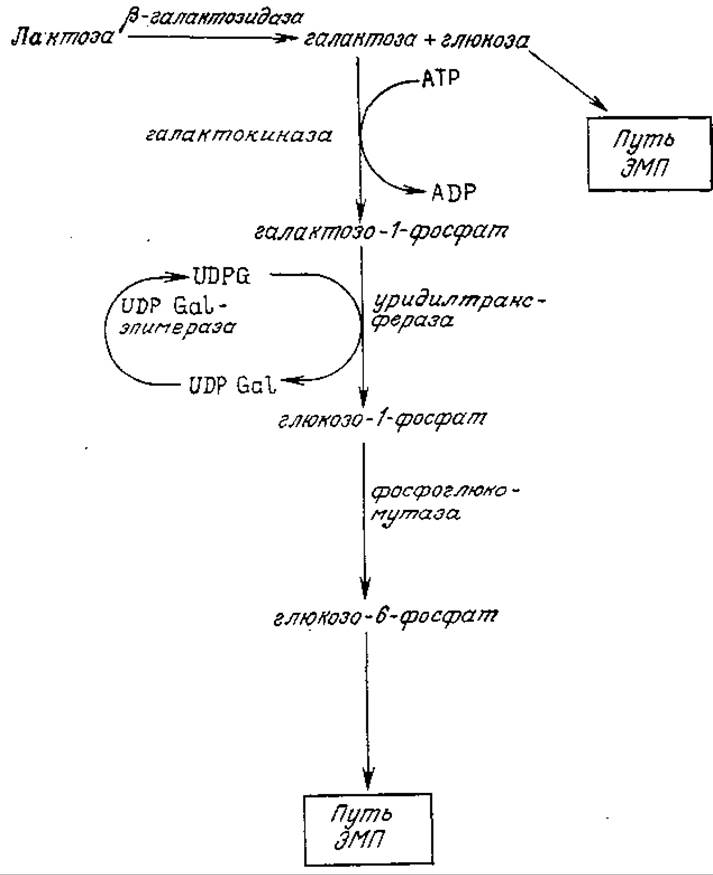
РИС. 5.5. Катаболизм лактозы Е. coli (UDPG — уридиндифосфатглюкоза; UDPGal — уридиндифосфатгалактоза).
Чрезвычайно интересны и разнообразны процессы утилизации микроорганизмами и других органических соединений, в частности углеводородов; любопытны и происходящие при этом химические превращения. Здесь мы ограничимся упоминанием о существовании в мире микроорганизмов ферментных систем, способных окислять углеводороды с изменением или без изменения их углеродного скелета, превращать углеводородные субстраты в аминокислоты, липиды, витамины, другие компоненты микробных клеток и иные соединения. Более того, некоторые микроорганизмы способны расщеплять токсичные вещества, например фенолы и полихлорбифенилы. Очевидно, такие микроорганизмы и синтезируемые ими ферменты потенциально очень важны как катализаторы специфических химических превращений, которые могут оказаться полезными в решении ряда проблем охраны окружающей среды. Введение в биохимию путей метаболизма углеводородов и соответствующих ферментов дано в превосходной книге Доуэлла [3].
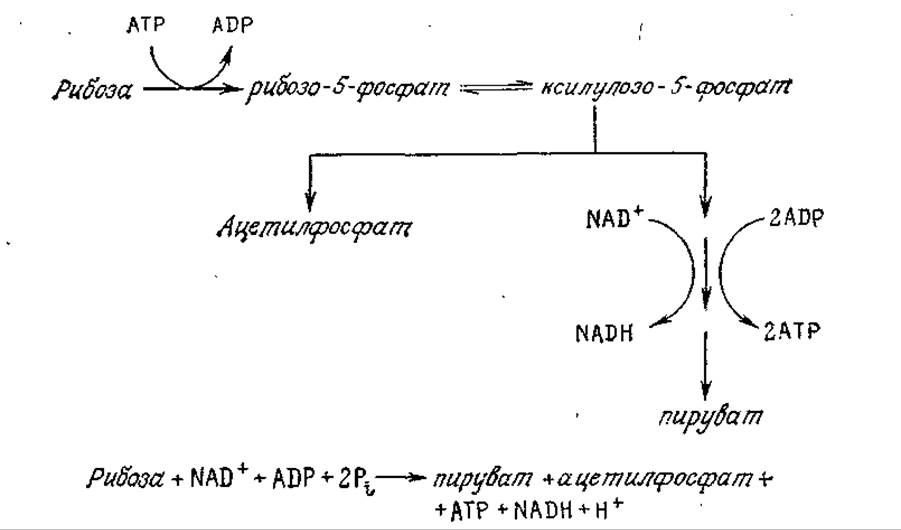
РИС. 5.6. Упрощенная схема катаболизма рибозы по фосфокетолазному пути.