Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Цикл лимонной кислоты
Как родилась сама мысль о существовании цикла лимонной кислоты?
Это законный вопрос, поскольку подобный цикл - с окислением двухуглеродных ацетильных групп до СО2 через шестиуглеродную лимонную кислоту - может показаться излишне сложным и следовательно, противоречащим принципу максимальной экономии, заложенному в биохимическую логику живой клетки.
Впервые предположение о существовании такого цикла для окисления пирувата в животных тканях было высказано в 1937 г. Гансом Кребсом. Эта идея родилась у него, когда он исследовал влияние анионов различных органических кислот на скорость поглощения кислорода суспензиями измельченных грудных мышц голубя, в которых происходило окисление пирувата. Грудные мышцы отличаются чрезвычайно высокой интенсивностью дыхания, что делает их особенно удобным объектом для изучения окислительной активности. Незадолго до описываемых работ Кребса Альберт Сент-Дьёрдьи в Венгрии обнаружил, что некоторые четырехуглеродные дикарбоновые органические кислоты, присутствующие в животных тканях (янтарная, фумаровая, яблочная и щавелевоуксусная), способны усиливать поглощение кислорода мышечной тканью. Кребс подтвердил это наблюдение и показал, что перечисленные органические кислоты стимулируют также окисление пирувата. Кроме того, он нашел, что окисление пирувата мышечной тканью стимулируется шестиуглеродными трикарбоновыми кислотами - лимонной, цис-аконитовой и изолимонной, а также пятиуглеродной к-кетоглутаровой кислотой. Структура всех этих кислот представлена на рис. 16-7. Испытаны были и некоторые другие встречающиеся в природе органические кислоты, но ни одна из них не обнаружила подобной активности. Обращал на себя внимание сам характер стимулирующего действия активных кислот: даже малого количества любой из них было достаточно для того, чтобы вызвать окисление во много раз большего количества пирувата.
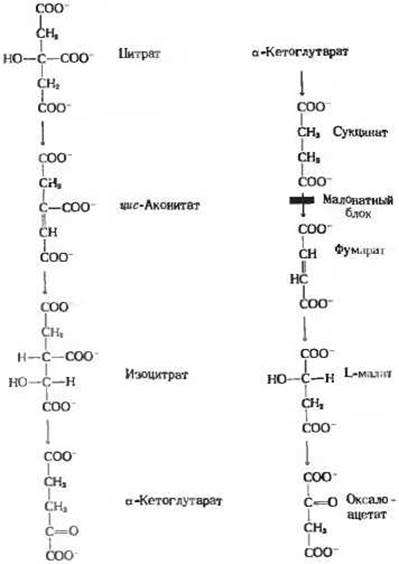
Рис. 16-7. Природные трикарбоновые и дикарбоновые кислоты, способные стимулировать окисление пирувата в суспензиях мышечной ткани. Другие встречающиеся в природе органические кислоты, например виннокаменная, щавелевая и кетоадипиновая, такой способностью не обладают. Активные кислоты приведены здесь в той последовательности, в какой они появляются в цикле лимонной кислоты. На каждом этапе происходит только одно химическое изменение. Отмечен этап, ингибируемый малонатом: в присутствии малоната цитрат окисляется до сукцината, который накапливается.
Другое важное наблюдение Кребса касалось действия малоната (рис. 16-8), одного из конкурентных ингибиторов сукцинатдегидрогеназы (разд. 9.13). Выяснилось, что малонат ингибирует аэробное потребление пирувата суспензиями измельченной мышечной ткани независимо от того, какая из активных кислот добавлена к суспензии. Это свидетельствовало о том, что сукцинат и сукцинатдегидрогеназа составляют необходимое звено в цепи ферментативных реакций, из которых слагается окисление пирувата. Кребс нашел далее, что если ингибировать малонатом аэробное потребление пирувата суспензией мышечной ткани, то в суспендирующей среде будут накапливаться цитрат, а-кетоглутарат и сукцинат. Отсюда следовало, что в норме, т.е. в отсутствие малоната, цитрат и а-кетоглутарат превращаются в сукцинат.
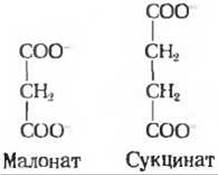
Рис. 16-8. Малонат - один из конкурентных ингибиторов сукцинатдегидрогеназы (см. также рис. 19-12). Обратите внимание на сходство структуры малоната и сукцината.
Исходя из этих основных данных и из некоторых других наблюдений, Кребс пришел к выводу, что все перечисленные выше активные три- и дикарбоновые кислоты можно расположить в некой логической химической последовательности, каждый этап которой представляет собой простое химическое превращение, катализируемое одним специфичным ферментом (рис. 16-7). Далее, поскольку инкубация пирувата и оксалоацетата с измельченной мышечной тканью приводила к накоплению цитрата в среде, Кребс сделал вывод, что эта последовательность является не линейной, а циклической, т.е. что ее начало смыкается с ее концом (рис. 16-9). Недостающим звеном, замыкающим цикл, должна была служить реакция
Пируват + Оксалоацетат →
→ Цитрат + СО2.
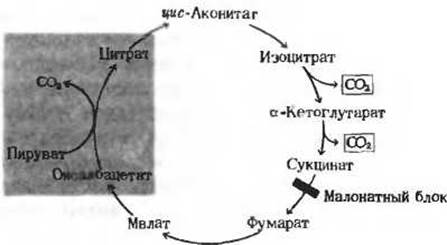
Рис. 16-9. Замыкание цикла в первой из предложенных схем. Когда Кребс обнаружил, что пируват и оксалоацетат взаимодействуют с образованием питрата (реакции на красном фоне), то стало ясно, что последовательность реакций имеет циклический характер. Обратите внимание, что сукцинат накапливается при малонатном блоке, когда пируват и оксалоацетат окисляются через цитрат.
Описанные выше простые эксперименты, а также логические рассуждения позволили Кребсу высказать предположение, что цикл, который он назвал циклом лимонной кислоты, является главным путем окисления углеводов в мышце. За годы, прошедшие со времени открытия этого цикла, выяснилось, что он существует не только в мышцах. Цикл лимонной кислоты обнаружен практически во всех тканях высших животных и растений и у многих аэробных микроорганизмов. За это важное открытие Кребс был удостоен в 1953 г. Нобелевской премии, которую он разделил с Фрицем Липманом, «отцом» АТР-цикла (разд. 14.7).
Цикл лимонной кислоты называют также циклом трикарбоновых кислот - это второе название появилось в связи с тем, что в течение нескольких лет после того, как Кребс постулировал существование цикла, не было полной уверенности в том, какая именно из трикарбоновых кислот (лимонная или, например, изолимонная) является первым продуктом конденсации пирувата с оксалоацетатом. Эта неопределенность, как мы увидим ниже, теперь устранена. В настоящее время точно известно, что первой из трикарбоновых кислот образуется именно лимонная кислота. Поэтому лучше всего называть данный метаболический путь циклом лимонной кислоты или просто циклом Кребса.

Рис. 16-10. Фотография сэра Ганса Кребса, сделанная по случаю его восьмидесятилетия в августе 1980 г. Кребс родился в Германии и там же получил медицинское образование. С 1926 по 1930 г. работал в Берлине с Отто Варбургом, который сам был одним из основоположников современной биохимии. В 1932 г., выполняя обязанности ассистента на медицинском факультете Фрейбургского университета, Кребс совместно с одним из студентов факультета Куртом Хенсейлайтом разработал схему постулированного ими цикла мочевины (гл. 19). В 1933 г. Кребс эмигрировал в Англию и начал работать в Кембриджском университете. Позже он перешел в Шеффилдский университет, где им и была выполнена большая часть работ по циклу лимонной кислоты. В 1954 г. он возглавил биохимический факультет в Оксфорде. После ухода с этого поста в 1967 г. Кребс вновь целиком посвятил себя исследовательской работе на медицинском факультете в Оксфорде. Он занялся изучением динамики и регуляции метаболизма и работал здесь очень активно вместе с несколькими сотрудниками вплоть до своей смерти в ноябре 1981 г. Кребс часто выступал с лекциями в университетах всего мира, и лекции его пользовались большой популярностью. Открытие цикла лимонной кислоты считается одним из важнейших в истории биохимии метаболизма.
Юджин Кеннеди и Альберт Ленинджер показали позднее, что все реакции цикла лимонной кислоты протекают в митохондриях животных клеток. В изолированных митохондриях печени крысы (разд. 2.8) были обнаружены не только все ферменты и коферменты цикла лимонной кислоты; здесь же, как выяснилось, локализованы все ферменты и белки, которые требуются для последней стадии дыхания, т. е. для переноса электронов и окислительного фосфорилирования. Поэтому митохондрии с полным правом называют «силовыми станциями» клетки. На рис. 16-11 показано, как размещены в митохондриях ферменты, катализирующие реакции цикла лимонной кислоты.
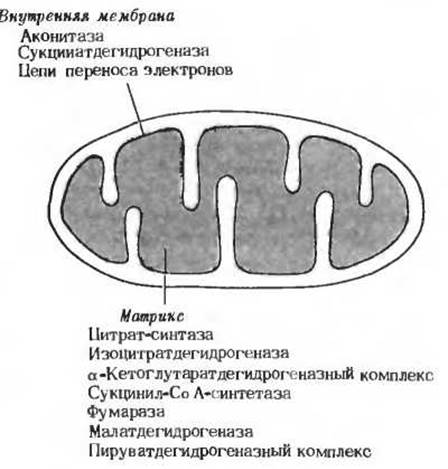
Рис. 16-11. Локализация ферментов цикла лимонной кислоты в митохондриях.