Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Перенос электронов, окислительное фосфорилирование и регуляция синтеза АТР
Согласно хемиосмотической гипотезе энергия переноса электронов передается на синтез АТР через протонный градиент
Именно те свойства митохондрий, которые мы только что рассмотрели, послужили основой для разработки хемиосмотической гипотезы (рис. 17-19). Согласно этой гипотезе, функция переноса электронов, происходящего во внутренней митохондриальной мембране, заключается в том, чтобы откачивать ионы Н+ из матрикса митохондрии в наружную среду и таким путем создавать между двумя водными фазами, которые разделяет эта мембрана, градиент концентрации ионов Н+ с более кислым значением pH снаружи. Такой градиент, при котором концентрация ионов Н+ снаружи выше, чем внутри митохондрии, обладает потенциальной энергией (разд. 14.16). Хемиосмотическая гипотеза постулирует далее, что ионы Н+, выведенные наружу за счет энергии переноса электронов, снова устремляются внутрь, в митохондриальный матрикс, через специальные каналы, или «поры», для этих ионов в молекулах F0F1-ATPaзы. В этом случае они перемещаются по градиенту концентрации и во время их перехода через молекулы АТРазы выделяется свободная энергия. Именно эта энергия и служит движущей силой для сопряженного синтеза АТР из ADP и фосфата.
Итак, хемиосмотическая гипотеза не требует никакого высокоэнергетического химического агента, который в качестве общего промежуточного продукта обеспечивал бы сопряжение между переносом электронов и синтезом АТР. Переносчиком энергии, связывающим два эти процесса, служит, согласно хемиосмотической гипотезе, градиент концентрации ионов Н+ между двумя сторонами митохондриальной мембраны. В свете этой гипотезы становится понятным и требование целостности мембраны, т. е. полной ее замкнутости в интактных митохондриях или в мембранных пузырьках, образовавшихся из разрушенной внутренней мембраны (рис. 17-15); ясно, что без этой целостности градиент концентрации ионов Н+ между двумя сторонами мембраны попросту не мог бы существовать. Нетрудно видеть также, что при «утечке» ионов Н+ через мембрану в присутствии разобщающих агентов (рис. 17-17) Н+-градиент должен «разряжаться». т. е. энергетическое сопряжение должно ослабевать. Наконец, удалось показать, что выведение ионов Н+ из митохондрий во время переноса электронов и поглощение наружных ионов Н+ молекулами АТР-синтетазы сопоставимы по своей скорости с процессом окислительного фосфорилирования в интактных митохондриях.
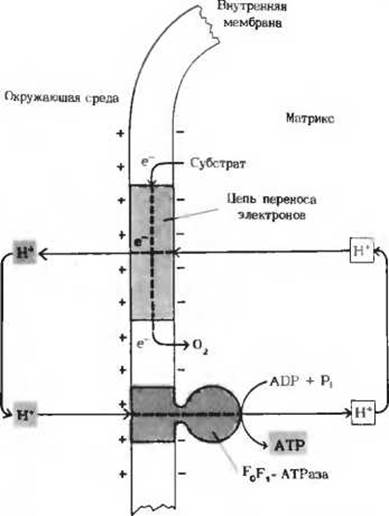
Рис. 17-19. Сопряжение переноса электронов с синтезом АТР в свете хемиосмотической гипотезы. Согласно этой гипотезе, цепь переноса электронов можно представить себе как насос, перекачивающий ионы Н+. Энергия, высвобождаемая при переносе электронов, используется для перемещения ионов Н+ из митохондриального матрикса наружу, что приводит к возникновению электрохимического Н+-градиента с более высокой концентрацией ионов Н+ в наружной водной фазе. Этот же процесс ведет к появлению трансмембранного электрического потенциала наружная сторона мембраны оказывается электроположительной.
Ионы Н+ из окружающей среды вновь устремляются внутрь, т.е. в митохондриальный матрикс, на этот раз по электрохимическому градиенту через молекулы F0F1-АТРазы. Этот переход ионов Н+ из зоны с более высокой в зону с более низкой их концентрацией сопровождается выделением свободной энергии, за счет которой и синтезируется АТР.
Таким образом, хемиосмотическая гипотеза предполагает, что между митохондрией и окружающей ее средой совершается непрерывный круговорот ионов Н+, движущей силой которого являемся перенос электронов (см. дополнение 17-1).
Однако хотя хемиосмотическая гипотеза удовлетворительно объясняет большую часть особенностей окислительного фосфорилирования, есть и такие стороны этого процесса, которые все еще остаются неясными. Загадочным остается, в частности, механизм, с помощью которого цепь переноса электронов «откачивает» ионы Н+ из матрикса митохондрий наружу (дополнение 17-1).
Дополнение 17-1. Многое в механизме окислительного фосфорилирования остается еще неясным
Хотя хемиосмотическая гипотеза получила широкое признание в той своей части, которая касается главного организующего принципа передачи энергии от процесса переноса электронов к синтезу АТР в митохондриях, бактериальных клетках и хлоропластах (гл. 23), тем не менее она оставляет пока без ответа многие важные вопросы. Пожалуй, больше всего споров порождает вопрос о механизме, при помощи которого перенос электронов, происходящий во внутренней мембране, вызывает откачивание ионов Н+ из матрикса митохондрии наружу. Митчелл предложил остроумное решение этого вопроса (рис. 1). Основой его решения послужил тот факт, что восстановительные эквиваленты переносятся некоторыми переносчиками (например, убихиноном) в виде атомов Н. а другими (например, железо-серными центрами или цитохромами) - в виде электронов. Митчелл предположил, что водородпереносящие и электронпереносящие белки чередуются в дыхательной цепи, образуя в ней три «петли». В каждой такой петле два атома Н выносятся через мембрану наружу и отдают два иона Н+ в окружающую среду; соответствующая пара электронов переносится затем обратно, с наружной поверхности мембраны на внутреннюю (рис. 1). Каждая пара восстановительных эквивалентов, проходя через такую петлю, переносит два иона Н+ из матрикса в окружающую среду. Предполагается, что каждая петля поставляет осмотическую энергию для образования одной молекулы АТР.
Этот гипотетический механизм представляется достаточно привлекательным, однако он подтверждается не всеми экспериментальными данными. Во-первых, он постулирует определенную последовательность расположения и чередование водород- и электронпереносящих центров, а это не вполне согласуется с имеющимися данными. Во-вторых, при таком механизме на каждую пару электронов в одной петле могут переноситься только два иона Н+, поскольку каждый из выходящих наружу электронов сопровождается только одним протоном. Между тем недавние исследования показали, что на каждую пару электронов в одной петле переносится не менее трех, а возможно, и четыре иона Н+ и что на каждую синтезированную молекулу АТР три или четыре иона Н+ возвращаются в матрикс.
Возникают и другие вопросы. Действительно ли нормальный процесс окислительного фосфорилирования сопровождается выходом ионов Н+ из митохондрий и их возвращением в матрикс? Складывается впечатление, что по крайней мере какая-то часть этого перемещения ионов Н+ может происходить внутри мембраны или же на ее поверхности, а не между двумя водными фазами, разделенными мембраной. Остается без ответа и вопрос о том, каким же именно образом поток ионов Н+ через сложную АТР-синтетазную систему создает новую ковалентную связь, посредством которой присоединяется концевая фосфатная группа АТР.
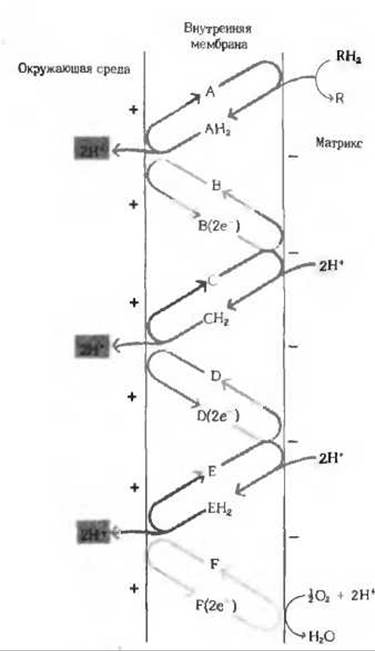
Рис. 1. Механизм переноса ионов Н+, постулируемый хемиосмотической гипотезой. Предполагается. что последовательные переносчики дыхательной цепи (А—F) образуют три Н+- переносящие «петли». Каждая такая петля переносит из митохондриального матрикса наружу два иона Н+ через переносчик (красные стрелки), транспортирующий восстановительные эквиваленты в виде атомов водорода.
Два электрона, оставшиеся после выведения в среду двух ионов Н+, возвращаются обратно, т.е. переходят на другую сторону мембраны, с помощью переносчика (серые стрелки), транспортирующего восстановительные эквиваленты в виде электронов. На каждую пару электронов, поступающих от субстрата RH2 на кислород, эти три петли переносят из митохондриального матрикса в среду шесть ионов водорода (3∙2 = 6Н+). Предполагается, что все компоненты дыхательной цепи фиксированы на мембране. Этим обеспечивается их необходимое расположение друг относительно друга.
Итак, нам предстоит выяснить еще очень многое о молекулярных компонентах и о свойствах энергопреобразующих мембран в митохондриях, бактериальных клетках и хлоропластах. Когда-нибудь, после проведения многих экспериментов и проверки новых идей, мы получим ответы на эти вопросы, но пока их у нас еще нет и виной тому в значительной мере сложная структура внутренней мембраны. Таков путь научного поиска: исследователи строят свои гипотезы, отталкиваясь от экспериментальных наблюдений, а затем проверяют их вновь и вновь, чтобы удостовериться в том, что ни один обнаруженный факт не остался без надлежащего объяснения. В известном смысле можно сказать, что биологическое исследование по-настоящему никогда не кончается. Нередко то, что представлялось нам твердо установленным, оказывается всего лишь неким приближением к истине, только шагом на пути к лучшему пониманию, открывающемуся с появлением новых фактов и новых представлений. Исследование молекулярной логики живых клеток - поистине безграничная область.