Основы биохимии Том 2 - А. Ленинджер 1985
Биоэнергетика и метаболизм
Перенос электронов, окислительное фосфорилирование и регуляция синтеза АТР
В окислении внемитохондриального NADH участвуют челночные системы
NADH-дегидрогеназа внутренней митохондриальной мембраны может присоединять электроны только от NADH, находящегося в матриксе. Внутренняя митохондриальная мембрана непроницаема для наружного NADH, который находится в цитозоле. Каким же образом может NADH, образующийся в процессе гликолиза, который, как известно, протекает вне митохондрий, вновь окисляться с образованием NAD+ молекулярным кислородом через дыхательную цепь?
Оказывается, существуют особые челночные системы, переносящие восстановительные эквиваленты от цитозольного NADH в митохондрии непрямым путем. Самая активная из них - это так называемая малат-аспартатная челночная система, действующая в митохондриях печени, почек и сердца. Рис. 17-26 поясняет принцип функционирования этой системы. От цитозольного NADH восстановительные эквиваленты сначала переносятся цитозольной малатдегидрогеназой на цитозольный оксалоацетат, что приводит к образованию малата. Малат, несущий восстановительные эквиваленты, полученные от цитозольного NADH, проходит через внутреннюю мембрану митохондрии в матрикс - его переносит через мембрану система, транспортирующая дикарбоксилаты. Попав внутрь митохондрии, малат отдает эти восстановительные эквиваленты NAD+ матрикса в реакции, катализируемой матриксной малатдегидрогеназой. NAD+ восстанавливается при этом в NADH, который может теперь передавать свои электроны прямо в дыхательную цепь внутренней митохондриальной мембраны. На каждую пару электронов, переданных на кислород, синтезируются три молекулы АТР. Другие компоненты этой челночной системы (рис. 17-26) регенерируют цитозольный оксалоацетат; это необходимо для того, чтобы мог начаться новый оборот челночного цикла.
В скелетных мышцах и в мозге перенос восстановительных эквивалентов от NADH осуществляется челночной системой другого типа. Это так называемая глицеролфоефатная челночная система. Она отличается от описанной выше малат-аспартатной челночной системы конечным этапом своего действия. Отличие состоит в том, что восстановительные эквиваленты передаются ею в дыхательную цепь не на участке 1, а на участке 2. Окисление NADH дает в этом случае не три молекулы АТР, а только две.
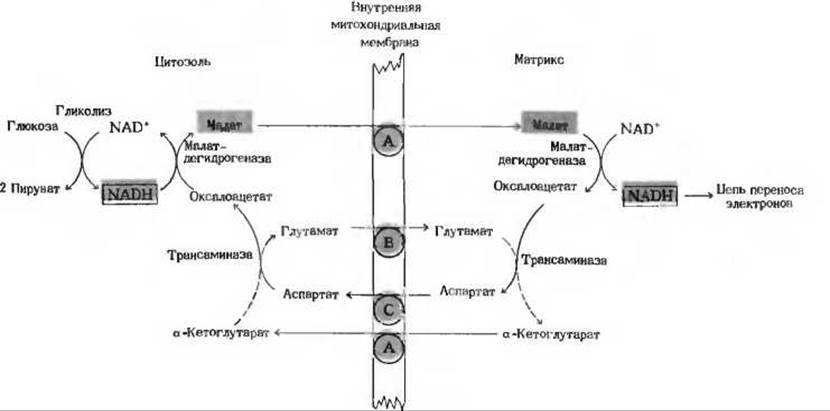
Рис. 17-26. Малат-аспартатная челночная система для переноса восстановительных эквивалентов от цитозольного NADH в митохондриальный матрикс. Несущий восстановительные эквиваленты малат переносится через внутреннюю мембрану при помощи системы, транспортирующей дикарбоксилаты (А). Затем эти восстановительные эквиваленты передаются на матриксный NAD+ малатдегидрогеназой, находящейся в матриксе. Образовавшийся в результате этого уже в матриксе NADH окисляется митохондриальной цепью переноса электронов с одновременным окислительным фосфорилированием. Продукт малатдегидрогеназной реакции, оксалоацетат, не способен пройти через мембрану, чтобы возвратиться в цитозоль. Под действием трансаминазы он превращается в аспартат, который может быть перенесен через мембрану системой С, транспортирующей аминокислоты. Назначение других реакций и транспортной системы В заключается в том, чтобы регенерировать в цитозоле оксалоацетат. Транспортная система В обеспечивает возможность обмена глутамата на аспартат. Система А переносит а-кетоглутарат наружу в обмен на малат, поступающий внутрь. Трансаминаза (гл. 19) катализирует обратимый перенос аминогрупп от глутамата на оксалоацетат:
Глутамат + Оксалоацетат ⇄ а-Кетоглутарат + Аспартат.