Биохимия - Химические реакции в живой клетке Том 3 - Д. Мецлер 1980
Метаболизм азотсодержащих соединений
Включение NH3 в аминокислоты и белки
До 1940 г. аминокислоты обычно рассматривались как относительно стойкие строительные блоки, поступающие в организм с пищей. От этих представлений быстро отказались после начатых Шёнкеймером исследований метаболизма 15NH3 и аминокислот, меченных изотопом 15N. Сразу же обнаружилось, что азот часто быстро переходит из одного углеродного остова в другой. Эти результаты подтвердили предположения, выдвинутые ранее Браунштейном (гл. 8, разд. Д). Браунштейн указывал, что С4- и C5-аминокислоты, аспартат и глутамат, тесно связанные с циклом трикарбоновых кислот, способны быстро обменивать свои аминогруппы на аминогруппы других аминокислот путем переаминирования [уравнение (14-12), стадии б и в]. Поскольку при этом аммиак легко включается в глутамат [уравнение (14-12), стадия а; см. следующий раздел], нетрудно представить себе существование общего пути синтеза аминокислот.
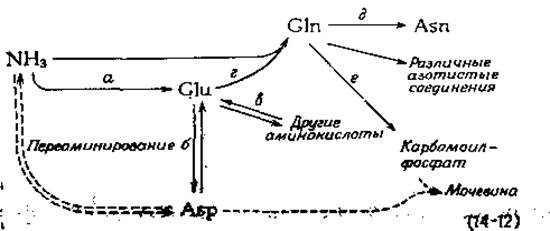
Вскоре стало ясно, что глутамин и аспарагин следует рассматривать как растворимые и нетоксичные переносчики дополнительного количества аммиака, заключенного в их амидных группах. Под действием активной синтетазы из глутамата и аммиака образуется глутамин [уравнение (14-12), стадия г], а под действием другого фермента происходит перенос амидного азота на аспартат с образованием аспарагина [уравнение (14-12), стадия д]. Амидный азот глутамина используется в многочисленных биохимических процессах, в том числе в образовании карбамоилфосфата [уравнение (14-12), стадия е; разд. В, 2], глюкозамина [уравнение (12-4)], NAD+ (разд. И), пуринов (разд. Л,3), СТР (разд. Л, 1), n-аминобензоата (разд. 3,3) и гистидина (разд. К).
Глутамат, глутамин и аспартат играют центральную роль н в удалении азота из органических соединений [17]. Будучи реакцией обратимой, переаминирование обычно служит начальным этапом катаболизма избыточных аминокислот. В результате присоединения азота к кето-глутарату образуется избыточный глутамат, который дезаминируется с образованием аммиака и далее — глутамина. Глутамин может также отдавать свой азот на образование аспартата. В организме животного и аспартат, и глутамин (через карбамоилфосфат) являются предшественниками мочевины, главного экскреторного азотистого соединения. Все эти взаимосвязи суммированы в уравнении (14-12), а дальнейшие подробности будут даны в последующих разделах.
Образование глутамата в результате восстановительного аминирования представляет собой основной путь включения азота в состав аминогрупп, однако вполне возможно, что существуют другие пути. Так, например, высказывалось предположение, что у растений происходит прямое аминирование пирувата и других а-оксокислот в ходе реакций, аналогичных реакции, катализируемой глутаматдегидрогеназой [17а]. Известен бактериальный фермент, который катализирует обратимое присоединение аммиака к фумарату с образованием аспартата (гл. 7, разд. 3,6, г).