БИОХИМИЯ - Л. Страйер - 1984
ТОМ 2
ЧАСТЬ II ГЕНЕРИРОВАНИЕ И ХРАНЕНИЕ МЕТАБОЛИЧЕСКОЙ ЭНЕРГИИ
ГЛАВА 13. ЦИКЛ ТРИКАБОНОВЫХ КИСЛОТ
13.6. При окислительном декарбоксилировании альфа-оксоглутарата образуется сукциннл-СоА
За превращением изоцитрата в а-оксоглу- тарат следует вторая реакция окислительного декарбоксилирования — образование сукцинил-СоА из а-оксоглутарата:
а-Оксоглутарат + NAD+ + СоА ⇄ Сукцинил-СоА + СO2 + NADH.
Реакция катализируется а-оксоглутарат- дегидрогеназным комплексом, организованным ансамблем, состоящим из трех видов ферментов. Механизм этой реакции очень сходен с механизмом превращения пиру- вата в ацетил-СоА. В обоих случаях используются одни и те же кофакторы: NAD+, СоА, тиаминпирофосфат, липоамид и FAD. Пируват-дегидрогеназный комплекс и α-оксоглутарат-дегидрогеназный комплекс имеют в действительности
ряд общих структурных особенностей (разд. 13.11).
13.7. Генерирование высокоэнергетической фосфатной связи из сукцинилкофермента А
В сукцинильном тиоэфире СоА имеется связь, богатая энергией. G0' для гидролиза сукцинил-СоА около — 8 ккал/моль, что сравнимо с этой величиной для АТР (- 7, 3 ккал/моль). Расщепление тиоэфирной связи сукцинил-СоА сопряжено с фосфорилированием гуанозиндифосфата (GDP):
Сукцинил-СоА + Рi + GDP ⇄ Сукцинат + GTP + СоА.
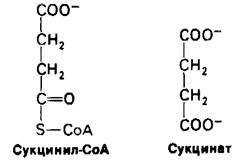
Эта легкообратимая реакция ∆G0' = - 0,8 ккал/моль) катализируется сукцинил-СоА—синтетазой. Фосфорильная группа гуанозинтрифосфата (GTP) в ходе реакции, катализируемой нуклеозиддифос- фокиназой, легко переносится на аденозиндифосфат (ADP) с образованием АТР.
GTP + ADP ⇄ CDP + АТР.
Образование высокоэнергетической фосфатной связи из сукцинил-СоА представляет собою пример субстратного фосфорилирования. В самом деле, это единственная реакция цикла трикарбоновых кислот, непосредственно приводящая к образованию высокоэнергетической фосфатной связи. По контрасту с нею, в процессе фосфорилирования, связанного с дыхательной цепью (называемого также окислительным фосфорилированием), образование АТР сопряжено с окислением NADH или FADH2 под действием 02. Мы уже встречались ранее с субстратным фосфорилированием при анализе двух реакций гликолиза: окисления глицеральдегид-3-фосфата и превращения фосфоенолпирувата в пируват. Окислительному фосфорилированию посвящена следующая глава.
13.8. Регенерирование оксалоацетата при окислении сукцината
Реакции соединений, содержащих четыре атома углерода, составляют последнюю стадию цикла трикарбоновых кислот (рис. 13.4). Сукцинат превращается в оксалоацетат в три этапа: окисление, присоединение молекулы воды (гидратация) и вторая реакция окисления. Таким образом, при каждом обороте цикла происходит регенерирование одной молекулы оксалоацетата и одновременно улавливание энергии в форме FADH2 и NADH.
Рис. 13.4. Последняя стадия цикла трикарбоновых кислот: от сукцината к оксалоацетату
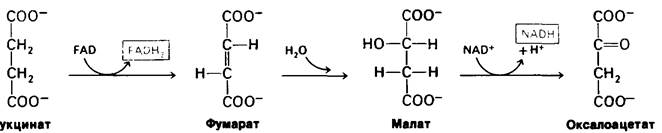
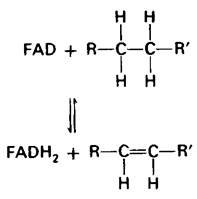
Сукцинат окисляется в фумарат сукцинат-дегидрогеназой. Акцептором водорода в этой реакции служит FAD, а не NAD+, который используется в трех других окислительных реакциях цикла. Роль акцептора водорода в этой реакции выполняет FAD, потому что изменение свободной энергии оказывается недостаточным для восстановления NAD+. В реакциях такого типа функцию акцептора электронов почти всегда несет FAD. В молекуле сукцинат-дегидрогеназы изоаллоксазиновое кольцо FAD ковалентно связано с боковой цепью гистидина, входящего в состав фермента (Е-FAD) .
Е — FAD + Сукцинат ⇄ Е — FADH2 + Фумарат.
Молекула сукцинат-дегидрогеназы содержит кроме флавина четыре атома железа и четыре неорганических сульфида. Гем в состав этого фермента не входит. Атомы железа связаны с неорганическими сульфидами. Белки такого типа известны как железосеропротеины (Fe-S-белки), или белки, содержащие негемовое железо. Железосеропротеины играют важную роль в электронтранспортных системах митохондрий и хлоропластов (разд. 14.4 и 19.9). Сукцинат-дегидрогеназа состоит из компонента 70 кДа, включающего FAD и два кластера FeS-белков, и компонента 27 кДа, содержащего единичный кластер белков. Этот фермент в отличие от других ферментов цикла трикарбоновых кислот является интегральным белком внутренней мембраны митохондрий. Сукцинат-дегидрогеназа непосредственно связана с цепью переноса электронов. FADH2, образовавшийся при окислении сукцината, не отделяется от фермента (в противоположность NADH). Два электрона FADN2 прямо переносятся на атомы Fe3+ фермента. Конечным акцептором этих электронов служит молекулярный кислород, что будет рассмотрено в гл. 14.
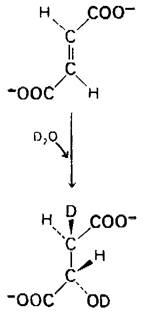
Следующий этап цикла-гидратация фумарата с образованием L-малата. Фумараза катализирует стереоспецифическое транс-присоединение Н и ОН, как было показано в исследованиях с использованием в качестве метки дейтерия. Поскольку ОН-группа присоединяется только с одной стороны двойной связи фумарата, образуется исключительно L-изомер малата.
В конце концов малат окисляется, образуя оксалоацетат. Эта реакция катализируется малат-дегидрогеназой, и роль акцептора электронов в ней опять выполняет NAD+.
Малат + NAD+ ⇄ Оксалоацетат + NADH + Н+.