БІОТЕХНОЛОГІЯ - В. Г. Герасименко - 2006
Частина І. Загальна біотехнологія
РОЗДІЛ 4. КЛІТИННА ІНЖЕНЕРІЯ
4.3.БІОТЕХНОЛОГІЯ ТРАНСПЛАНТАЦІЇ ЯДЕР
Більшість досліджень, виконаних в останні роки з гібридизації соматичних клітин еукаріотичних організмів (переважно ссавців) у клітинній культурі, переконливо довели, що в гібридній клітині, які має змішаний геном у зв’язку з гетерогенною природою батьківських клітин, відбуваються стабільні зміни в експресії генів синкаріона порівняно з процесом експресії у вихідних батьківських клітинах. Так, при злитті клітин щурячої гепатоми, що синтезує і екскретує альбумін, з мишачими фібробластами, що не секретують цей білок, були отримані гібридні клітини, частина клонів яких синтезували альбумін тільки пацюка, чи тільки миші, або альбуміни обох вихідних батьків (пацюка і миші).
На підставі результатів цього й інших подібних експериментів з гібридизації соматичних клітин було висловлене припущення про те, що в клітинах ссавців є речовини, здатні прямо чи побічно впливати на клітинне ядро, змінювати його функціональний стан за допомогою позитивної або негативної регуляції експресії певної частини генів. Однак комплекс об’єктивних причин (гетерогенність генома найчастіше зі значним дефіцитом хромосом, а також із хромосомними перебудовами, що відбулися, які знаходяться в оточенні гетерогенної цитоплазми), властивих методу гібридизації соматичних клітин, не дозволив ідентифікувати речовини, що беруть участь в експресії генома, а також установити їхню хімічну будову і механізм дії. До цього часу вчені мали у своєму розпорядженні дані про те, що речовини, які містяться в цитоплазмі тварин, беруть участь у регулюванні експресії генів клітинного ядра.
Виявилося, що в ядрах попередньо вилучених з диференційованої клітини і поміщених у цитоплазму яйцеклітини починалися процеси клітинного диференціювання і в багатьох випадках розвивався нормальний організм, тобто ядра спеціалізованих клітин містили повний обсяг необхідної для розвитку повноцінного організму інформації. Так, злиття ядер, вилучених з клітин кишкового епітелію пуголовка, з позбавленої ядра яйцеклітиною в багатьох випадках приводило до розвитку жаби. Ці експерименти, виконані на амфібіях, показали, що цитоплазма регулює активність ядра; у спеціалізованих клітинах багато функцій ядра гальмуються компонентами цитоплазми. Інгібуючий вплив, що надходив до ядра з боку цитоплазми диференційованої клітини, опинившись у придатному за хімічним складом цитоплазматичному оточенні (яйцеклітина), усувається, і в ядрах можуть знову проходити процеси, що визначають клітинну диференціацію (Gurdan J.B., 1977). Експерименти з трансплантації ядер були проведені на амфібіях. У розробці методів пересадки клітинних ядер ссавців істотну роль відіграло використання цитохалазинів — речовин, синтезованих грибами.
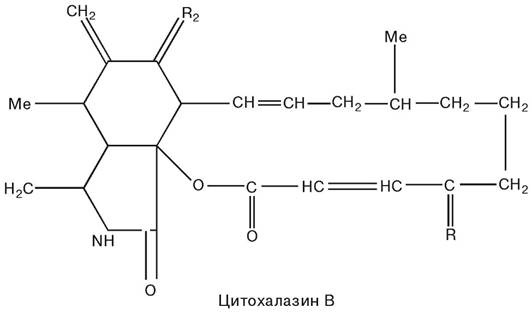
Висловлюється думка, що цитохалазин В, руйнуючи структуру мікрофіламентів, сприяє унікальному розташуванню ядра, коли воно залишається з’єднаним із клітиною тонкою стеблинкою цитоплазми. При центрифугуванні в більшості таких клітин розривається «пуповина» стеблинки й утворюються енуклейовані (без’ядерні) клітини (цитопласти); ядра, що відокремилися при центрифугуванні (каріопласти, чи міні-клітини), оточені тонким шаром цитоплазми і плазматичною мембраною.
Ефективність енуклеації контролювали, забарвлюючи (наприклад, за методом Гімза) клітинний моношар, що знаходиться на поверхні однієї з чашок або дисків.
Наступна операція, спрямована на одержання популяції цитопластів, — відділення їх від цілих клітин, що залишилися, тому що в деяких випадках (лінія клітин мишачої гепатоми Hepa-2) ефективність енуклеації не перевищує 50 %. З цією метою, енуклейовані (цитопласти) і цілі клітини, що містяться на поверхні пластикових чашок і скляних дисків, знімали, обробляли трипсином, внаслідок чого в 1 мл сольового розчину з фосфатним буфером містилося 106 цілих клітин і цитопластів. Клітинно-цитопластичну суміш нашаровували на 14 мл лінійного градієнта ренографіну-76, який знаходився в центрифужній пробірці, об’ємна концентрація якого змінювалася від 15 до 30 %, і центрифугували при 1000 g і температурі 25 оС протягом 5 хв. Чітко розмежовані шари цілих клітин і цитопластів видаляли з центрифужної пробірки і використовували за призначенням.
Отримані в градієнті щільності ренографіну-76 цитопласти не забарвлювалися трипановим синім, що слугувало контролем ефективності поділу. Вони зберігали здатність знову прикріплюватися до поверхні культуральної чашки і могли використовуватися надалі для реконструювання життєздатних клітин шляхом злиття з гетерологічними каріопластами.
Методи енуклеації, застосовані для одержання каріопластів, відрізняються від тих прийомів, що забезпечують успіх при одержанні цитопластів. Клітини, призначені для виділення каріопластів, за 2 дні до енуклеації висівають на вирізані з культуральних флаконів пластикові пластинки, які перед поміщенням у 50-мілілітрову центрифужну пробірку, заповнену звичайним поживним середовищем, складали так, щоб їхні поверхні з прикріпленими клітинами знаходилися зовні. Мишачі фібробластні лінії А9 центрифугували при 9,5 тис. g і 35 оС протягом 15 хв. Призначення цієї процедури зводилося до того, щоб в осаді каріопластів, який має бути отриманий, зменшити кількість цілих клітин. Після відокремлення слабко прикріплених до субстрату клітин, пластинки з клітинами моношару, що залишилися, переносили в пробірки, де концентрація цитохалазину В з розрахунку на 1 мл звичайного поживного середовища досягає 10 мкг. Вміст пробірок інкубували при 37 оС протягом 15 хв; тривалість центрифугування при 35 оС і 7 тис. об./хв — 45 хв. Надосадову рідину, що утворилася, зливали, а осад, представлений в основному каріопластами, ресуспендували в звичайному поживному середовищі.
У виготовлених за описаною методикою препаратах каріопластів знаходяться фрагменти цитоплазми, каріопласти, що загинули, та цілі клітини. Відокремлення фрагментів цитоплазми проводиться осадженням у градієнті фіколау 1-6% концентрації при 1 g і 37 оС протягом 90 хв у зволоженій атмосфері, що містить 5 % СО2. Фрагменти цитоплазми, що знаходяться у верхньому шарі, відсмоктують і видаляють, а осад каріопластів, що утворився після вилучення і розведення поживним середовищем, знову осаджують центрифугуванням і ресуспендують у свіжому живильному середовищі. Цілі клітини (їх 0,4-4 %) видаляють шляхом дворазового 90-хвилинного інкубування суспензії каріопластів у культуральних чашках.
Інкубування протягом 3 годин дозволяє різко зменшити забруднення каріопластів цілими клітинами, що обов’язково для усіх експериментів з трансплантацією ядер.
Наступна операція — відокремлення життєздатних каріопластів від тих, що загинули. Вона включає ресуспендування осаду в поживному середовищі до концентрації 107 міні-клітин у 1 мл, нашаровування суспензії на Ficollpaque (розчин містить 5 % фіколу і 9 % діатризоату натрію), що знаходиться в центри- фужних пробірках, і обережне додавання зверху 1 мл середовища так, щоб додане середовище і суспензія каріопластів утворили два самостійних шари. Наступне центрифугування при 800 об./хв (130 g) протягом 75 хв при кімнатній температурі дозволяє одержати осад, що на 99 % складається із загиблих каріопластів; на межі живильного середовища і Ficoll-paque знаходиться шар, що складається на 98 % з життєздатних каріопластів.
Для очищення каріопластів використовують й інші методи, зокрема частки танталу розміром 1-3 мкм. До часу енуклеації практично всі клітини містять більш ніж по 12 часток танталу. У зв’язку з тим, що щільність танталу більш ніж у 15 разів перевершує щільність клітини, компоненти, що містять частки танталу, осаджуються значно швидше каріопластів, у яких тантал відсутній. Очищення каріопластів іноді здійснюється за допомогою цитофлуориметра-сортера.
Властивості цитопластів і каріопластів. Установлено здатність цитопластів синтезувати білки, підтримувати реплікацію вірусу везикулярного стоматиту і поліовіруса, синтез РНК і білка, контрольований вірусом сказу, звільняти вірус SV40 13 трансформованих клітин. Цитопласти містять усі види органел, властиві нормальній клітині, зберігають характерну для цілих клітин здатність прикріплюватися до субстрату й утворювати складчасту мембрану, пересуватися і здійснювати піноцитоз.
Навколо каріопластів знаходиться шар, на частку якого припадає близько 10 % клітинної цитоплазми, що містить компоненти ендоплазматичного ретикулума, деяка кількість мітохондрій і рибосом; центріолі в каріопластів, на відміну від цитопластів, відсутні.
Близько 10 % каріопластів деяких клітинних ліній здатні відновлювати весь обсяг утраченої при енуклеації цитоплазми і знову перетворюватися на життєздатні клітини.
Здатність каріопластів регенерувати втрачену в процесі енуклеації цитоплазму і формувати життєздатні колонії клітин залежить від кількості цитоплазми, що оточує ядро. У фракції каріопластів, біля ядер яких зосереджено 2-4 % тієї кількості цитоплазми, що міститься в інтактній клітині (дрібні каріопла- сти), тільки один з 106 очищених елементів може сформувати життєздатну колонію клітин.
Трансплантація ядер і реконструювання клітин.
Після енуклеації клітин моношар цитопластів у пластикових чашках діаметром 60 мм чи скляних дисках діаметром
14 мм інкубують у поживному середовищі при 37 оС протягом 1-2 годин, а потім — 20 хв і охолоджують до 4 оС. При цій температурі моношар двічі відмивають розчином Ерла (рН 8,0), потім 20 хв. обробляють 0,5 мл охолодженого розчину Ерла з вірусом Сендай, інактивованим опроміненням протягом 5 хв (ультрафіолетовою лампою на відстані 15 см), після чого шар цитопластів знову двічі промивають тим же розчином, який потім видаляють.
Каріопласти, суспендовані в сольовому розчині на фосфатному буфері, додають до моношарової культури цитопластів з таким розрахунком, щоб на один цитопласт припадало 100 каріопластів. Для адсорбування каріопластів на цитопластах, покритих вірусними частками, інкубують при 4 оС протягом 45 хв; через кожні 3-5 хв чашки злегка погойдують. Для злиття каріопластів і цитопластів чашки чи скляні диски переносять у термостат і витримують там при 37 оС протягом 45 хв. Після закінчення цього часу моношар на чашках кілька разів інтенсивно відмивають розчином Ерла чи поживним середовищем без сироватки, щоб видалити каріопласти, які не злилися. Потім у чашки наливають поживне середовище, придатне для культивування того типу клітин, що були використані як донори каріопластів. Цитопласти, що не злилися з ядрами, гинуть і приблизно через 2 дні відокремлються від поверхні чашки.
Для одержання незабруднених цитопластами культур рекомендується центрифугування гібридних клітин у градієнті щільності ренографіну.
Використання поліетиленгліколю для стимулювання процесу злиття каріопластів і цитопластів при конструюванні клітинних гібридів обмежене через його токсичність, що значно перевищує токсичність вірусу Сендай.
Для ідентифікації гібридних клітин, визначення ефективності трансплантації ядер і наявності батьківських клітин у гібридній популяції використовують мутантні клітинні лінії.
Ефективність реконструювання клітин за рахунок злиття каріопластів і цитопластів залежить від багатьох факторів. Так, тільки близько 10 % цитопластів, отриманих шляхом енуклеації клітин пацюка лінії НТС, зливалися з каріопластами, виділеними з клітин миші лінії А9. Коли гібридизації піддавали цитопла- сти, отримані з фібробластів курячих ембріонів, і каріопласти, що являли собою перебуваючі у спокої ядра еритроцитів, ефективність реконструювання перевищувала 90 %. Висока ефективність злиття ізольованих перебуваючих у спокої ядер еритроцитів птахів з енуклейованими цитопластами, а також досвід щодо активування ядер еритроцитів птахів, зокрема курей, шляхом злиття цих еритроцитів із клітинами HeLa чи з іншими клітинами, що мають активний метаболізм, дозволили досліджувати роль ядерно-цитоплазматичних взаємодій в експресії генів гібридних еукаріотичних клітин. Виявилося, що не всі гібриди, що утворилися, були життєздатними. Приблизно 9 % реконструйованих клітин можуть рости і ділитися (Хайтауер М., Льюкас Дж., 1985). У гібридних клітинах відразу після злиття починаються морфологічні зміни. Через кілька днів реконструйовані гібридні клітини майже не відрізняються від вихідних батьківських, які було використано як донори ядер.
Трансплантація ізольованих інтерфазних ядер, укладених у ліпідну оболонку, у соматичні клітини.
У літературі є дані про введення молекул, у тому числі ДНК, у клітини за допомогою «тіней» еритроцитів (Рекстейнер М., 1985) і ліпосом (Штраубингер Р., Папахаджопулос Д., 1985). Так, Сєровим О.Л. (1985) у лабораторії генетичних основ онтогенезу Інституту цитології і генетики СО АН СРСР була виконана експериментальна робота щодо включення в штучні мембрани ізольованих інтерфазних ядер з метою забезпечення збереження генетичного матеріалу від деградації та індукування процесу трансформації ізольованих ядер у реципієнтні клітини. За даними Сєрова О.Л., інтерфазні ядра, виділені з культури фібробластів норки, суспендували в багатокомпонентному буферному розчині, що включає різні концентрації KCl, NaCl, трис-HCl, ЕДТА, сахарози і спермідину.
Для створення штучної мембрани навколо ядер використовували фосфатидилетаноламін у концентрації 10-20 мг/мл і фосфатидилхолін, отриманий із жовтків курячих яєць, у концентрації 150-200 мг/мл органічного розчинника. Запропонований Сєровим О.Л. метод заснований на утворенні фосфолі- підної мембрани при проходженні ізольованих ядер через тришарову систему, що знаходиться в центрифуговій пробірці з розчином 1М сахарози, яка міститься на дні центрифугової пробірки, водним розчином 0,25 М сахарози, що формує верхній шар, і шаром органічного розчинника, що складається з хлороформу, діоксану і етилацетату в співвідношенні 0,25:0,23:0,51 і фосфатидилхоліну в концентрації 150-200 мг/мл зазначеного органічного розчинника, що займає в центрифуговій пробірці положення між верхнім і нижнім шарами сахарози.
На межі трикомпонентного органічного розчинника з верхнім сахарозним шаром молекули фосфатидилхоліну орієнтовані своїми гідрофільними кінцями убік водного шару; на межі органічного розчинника з нижнім сахарозним шаром молекули фосфоліпіду також будуть орієнтовані своїми полярними голів
ками до водяної фази більш щільного нижнього сахарозного шару. Під час центрифугування ядра при переході з водного (0,25 М сахарози) у шар органічного розчинника на межі поділу фаз зустрічаються з полярними гідрофільними голівками молекул фосфатидилхоліну, який формує одношарову ліпідну мембрану на поверхні інтерфазних ізольованих ядер. На межі органічного шару з більш щільним шаром 1М сахарози ядра, що вже мають на своїй поверхні одношарову ліпідну мембрану, орієнтовану гідрофобними кінцями назовні, зустрічаються із шаром молекул фосфатидилхоліну, гідрофобні кінці яких спрямовані убік органічної фази і взаємодіють з гідрофобними кінцями ліпідного моношару ядер, утворюючи при цьому другий шар мембрани на поверхні інтерфазних ядер.
Сформований ліпідний бішар міцно утримується на поверхні інтерфазних ядер і не змивається при багаторазових пере- осадженнях шляхом центрифугування через розчин 1М сахарози. За допомогою радіоавтографії і флуоресцентної мікроскопії було показано, що ліпідний бішар рівномірно розподіляється на поверхні ядра, формуючи додаткову мембрану чи мембрани (при зазначених умовах центрифугування вільні ліпосоми в ядерну фракцію потрапити не можуть).
У результаті додаткових експериментів було показано, що штучно створена мембрана не є досконалою (Сєров О.Л.). Такий висновок, зроблений на підставі виявлення виходу з ядер, оточених фосфатидилхоліновою мембраною, АТФ і проникнення в ці ядра екзогенної ДНКази. Одночасно було встановлено, що в обох випадках утворена мембрана блокувала транслокацію як макромолекул (ДНКаза), так і звичайних молекул порівняно невеликих розмірів (АТФ). Тим часом відомо, що природні ядерні мембрани проникливі навіть для полімерних молекул; штучна мембрана, створена з фосфатидилхоліну, зовсім непрониклива як для великих полярних молекул, так і для макромолекулярних структур.
У результаті проведених експериментів була висловлена думка, що фосфатидилхолін екзогенного походження формує мембранну структуру навколо ядра, у зв’язку з чим воно виявляється заключеним у ліпосому, яка, будучи додатковою мембраною, накладає певні обмеження на перенесення деяких
молекул у ядро і назад, але цілком ці процеси не припиняються. При введенні інтерфазних ядер норки, що оточені штучною мембраною, в культуру LMTK — клітин миші, які характеризуються дефіцитом ферменту тимідинкінази (ТК), установлене перенесення у реципієнтні мишачі клітини порівняно великих фрагментів ДНК із ядер норки, а більшість досліджених трансформаторів характеризувалися стабільним ТК+ фенотипом.
Пропонований метод перенесення генетичного матеріалу за допомогою інтерфазних ядер, що знаходяться у ліпосомі, завдяки високій частоті трансформації в порівнянні з методом перенесення генів у складі сумарної клітинної ДНК свідчить про можливу захисну функцію ядерних білків донора, що захищають ДНК від деградації, а також про участь цих білків в інтеграції донорського хроматину і хроматину реципієнтних клітин.
Біотехнологія одержання цибридів. Цибридами називаються продукти, що утворюються при злитті цілих клітин однієї мутантної лінії з цитоплазмою енуклейованих клітин іншої клітинної лінії. Для одержання цибридів проводили енуклеацію клітин, попередньо маркірованих флуоресціюючими гранулами зеленого кольору, що додаються в поживне середовище за допомогою цитохалазину В, концентрацію якого доводили до 5 мкг/мл середовища.
Іонгкинд І. та Веркерк А. (1985) позначили фібробласти людини флуоресцентними барвниками, інкубуючи проліферуючі клітинні культури протягом 2 днів у середовищі F10 Хема, що містить у 1 мл 2-107 гранул барвника, 100 мкг стрептоміцину і 100 одиниць пеніциліну, з додаванням 10 % сироватки плода корови. Під час інкубації клітини, що знаходяться у культурі, фагоцитували гранули; гранули прикріплені до зовнішньої поверхні клітинної мембрани, змивали сольовим розчином. Як показали результати проведених досліджень, маркірування флуоресціюючими гранулами сповільнює ріст клітин.
Енуклейовану суспензію клітин (цитопластів) забарвлюють також флуоресцентними бісимідазоловими барвниками фірми Hoechst; флуоресціюючі цитопласти виділяють методом сортування у потоці за допомогою клітинного сортера. Чистота фракції цитопластів досягала 99,7 %, а 90 % отриманих після сортування цитопластів мають здатність до включення 3Н-лейцина.
Маркіровані тим чи іншим способом цитопласти зливаються з цілими клітинами, також попередньо позначеними флуоресціюючим барвником, що відрізняється за кольором від барвника, який був використаний для позначення енуклейованих клітин (цитопластів); співвідношення цитопластів і цілих клітин при одержанні цибридів — 3:1. Як агент, індукуючий процес злиття цитопластів і цілих клітин, використовують поліетиленгліколь (ПЕГ 1000), диметилсульфоксид чи інактивований вірус Сендай. Продукти злиття (цибриди) мають двоколірну флуоресценцію. їх виділяють за допомогою потокового клітинного сортера FACS ІІ, оснащеного аргоновим лазером, при довжині хвилі 488 нм і постійному виході 100 мВт. Шланги сортера, через які здійснюється подача зразка, стерилізуються 70 %-ним етанолом, а несуча рідина для стерилізації пропускається через бактеріальні фільтри (Millipore, пори 0,22 мкм). Якщо в клітинній суспензії після злиття цитопластів і цілих клітин було багато грудок, їх диспергують, пропускаючи через наконечник (діаметр 70 мкм) клітинного сортера.
Вихід фракції цибридів після проведеного сортування визначають, зливаючи контрольні цитопласти, що є носіями гіпоксантифосфорибозилтрасферазної (ГФРТ+) активності, з фібробластами, отриманими від хворих на синдром Леша — Ніхана (ГФРТ). Через 20 годин після злиття клітини з двоколірною флуоресценцією (цибриди) при сортуванні зміщують на покривні скельця, інкубують 20 годин із 3Н-гіпоксантином, фіксують і досліджують радіоавтографічним методом. Включення гіпоксантину в цибридну фракцію показує, що за допомогою вірусу Сендай не менш 90 % клітин, які пройшли сортування, можна віднести до справжніх цибридів (ГФРТ+).
Для визначення активності ферментів і концентрації субстратів у надто малому об’ємі матеріалу (5-10 тис. цибридів) були розроблені мікрометоди, засновані на застосуванні флуориметрії, що дозволяють виявити мікрограмові кількості речовин у незначних кількостях ліофілізованих клітин. Ультрамікрометод, заснований на проведенні мікрофлуориметрії, дає можливість вимірювати активність ферментів навіть в поодиноких клітинах. За допомогою цих методів з’явилася можливість вивчати комплементацію ферментів у цибридів чи інших клітин, а також з’ясувати роль ядра і цитоплазми в механізмі комплементації.